| |
|
|
|
|
|
 SCIENCES DE LA VIE : Des phénotypes à différents niveaux d'organisation du vivant. SCIENCES DE LA VIE : Des phénotypes à différents niveaux d'organisation du vivant. |
Du génotype au phénotype, relations avec l'environnement.
Chapitre du cours |
Corrigé |
| La diversité des phénotypes |
PLAN DETAILLE :
Comment peut-on expliquer les phénotypes différents des individus ?
I. Le phénotype se définit à plusieurs échelles : de l'organisme à la molécule.
A. Le phénotype macroscopique.
Les différences observées au niveau du phénotype macroscopique existent-elles au niveau cellulaire ?
B. Le phénotype cellulaire.
Les différences observées au niveau du phénotype cellulaire existent-elles au niveau moléculaire ?
C. Le phénotype moléculaire.
Comment expliquer cette variation des phénotypes pour un même caractère ?
II. Les phénotypes alternatifs sont dus à des différences dans les protéines concernées.
Comment distingue-t-on les protéines différentes ?
A. Technique d'étude des protéines : Cas de l'hémoglobine.
1) L'électrophorèse.
2) La chromatographie.
Quels sont les liens de cause et conséquence entre les différents niveaux de phénotype ?
B. Liens entre les 3 niveaux du phénotype. |
| Les enzymes, protéines indispensables à la réalisation du phénotype |
|
| Les enzymes, protéines indispensables à la réalisation du phénotype |
|
| Synthèse des protéines |
Plan détaillé :
Intro : Maladie héréditaire – gène – protéine.
I. Les étapes essentielles pour passer d’un gène à une protéine.
A) La transcription
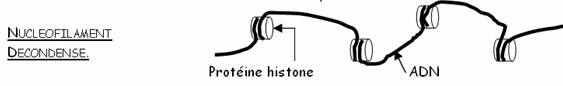
ADN –ARNn – Nucléotides noyau pores nucléaires – Autoradiographie + Schéma
Que devient cette petite molécule synthétisée dans le noyau ?
B) La traduction.
Code génétique – ribosome (sous unités) – acides aminés – polypeptides – liaisons peptidiques – structure primaire – séquences – cytoplasme – + Schéma
Comment le polypeptide devient-il protéine ?
C) La maturation
R.E.G. – Appareil de Golgi – vésicules – exocytose – conformation spatiale – structure quaternaire – fonction.
Comment expliquer que certaines personnes soient atteintes d’une maladie héréditaire comme la drépanocytose ?
II. Modification de la protéine suite à une mutation.
A) Les mutations.
Nucléotide – germinale – somatique – substitution – délétion – décalage du cadre de lecture
Quelles sont les conséquences des mutations ?
B) Conséquences des mutations sur les protéines.
1) Le cas de la drépanocytose.
HbA –HbS – Hémoglobine fibreuse – falciforme + Schéma
Toutes les mutations causent-elles des maladies génétiques ?
2) Des mutations sans conséquence.
Redondance- enzyme – site actif – conformation spatiale – fonction
Conclusion : Résumé - Phénotype cellulaire - Phénotype macroscopique
|
|
Synthèse des protéines
et
Complexité des relations entre gènes, phénotypes et environnement. |
|
| tous les chapitres de cette partie |
|
|
|
La régulation de la glycémie et les phénotypes diabétiques.
Chapitre du cours |
Corrigé |
| La régulation de la glycémie |
Sujet 1 : CORRIGE COMPLET (sauf schémas) :
La glycémie est une constance physiologique c’est-à-dire que sa valeur ne subit que des légères variations autour d’une valeur moyenne stable (1 g/l) ou notion du cours sur la variation de la glycémie au cours d’une journée.
Nous montrerons dans ce devoir comment est assuré le maintien dans l’organisme d’une valeur physiologique de la glycémie.
Pour cela nous présenterons les mécanismes assurant le stockage après un repas avant d’étudier ce qui se passe en cas de jeûne.
A. Mécanisme assurant le stockage du glucose en cas d’apport excessif.
1) Action des capteurs.
Les cellules bêta des îlots de Langerhans (schéma) :
- captent la concentration plasmatique du glucose (l’état d’hyperglycémie),
- sécrètent l’insuline qui est une hormone hypoglycémiante. La quantité d’insuline sécrétée dans le milieu constitue le message hormonal envoyé par les capteurs aux cellules effectrices pour réguler la glycémie.
2) Action des organes cibles.
Les cellules hépatiques et musculaires, les tissus adipeux et d’autres cellules possèdent les récepteurs spécifiques de l’insuline. La liaison hormone – récepteur entraîne la modification de l’activité de ces organes cibles :
- stockage sous forme de glycogène (glycogénogénèse) dans le foie et dans les muscles et sous forme de triglycérides dans les tissus adipeux (et dans le foie en cas d'excès important : foie gras !).
- et augmentation de l’utilisation du glucose par d’autres cellules entraînant ainsi le retrait du glucose du compartiment sanguin
B. Mécanisme assurant le déstockage du glucose.
1) Action des capteurs.
Les cellules alpha des îlots de Langerhans:
- captent la concentration plasmatique du glucose (l’état d’hypoglycémie),
- sécrètent le glucagon qui est une hormone hyperglycémiante. La quantité de glucagon sécrétée dans le milieu constitue le message hormonal envoyé par les capteurs aux cellules effectrices pour réguler la glycémie.
2) Action des organes cibles.
Les cellules hépatiques sont les seules cellules possédant les récepteurs spécifiques du glucagon. La liaison hormone – récepteur entraîne la modification de l’activité de cet organe cible : déstockage du glucose en hydrolysant le glycogène hépatique (glycogénolyse) entraînant ainsi le passage du glucose dans le compartiment sanguin. Le déstockage des triglycérides par néoglucogenèse est réalisé en cas de besoin important (et de réserves de graisses dans le foie).
Conclusion : + Schéma de la régulation de la glycémie.
Après un repas, le stockage et l’utilisation accrue du glucose est permis par l’action de l’insuline, synthétisée par les cellules β des îlots de Langerhans.
Au cours du jeûne, ce sont les cellules α qui agissent grâce au glucagon hypoglycémiant.
Comment peut-on alors expliquer que certaines personnes, qualifiées de diabétiques, ne parviennent pas maintenir la constance de leur glycémie ? |
| les phénotypes diabétiques |
Sujet 2
Introduction : Diabète de l’adulte – hyperglycémie – insulinorésistance - déficit (+ tardif) de l’insulino-sécrétion. Quelles sont les causes du diabète de type 2 et comment peut-on prévenir cette maladie ?
I. Facteurs à l’origine du diabète de type 2.
A. Facteurs génétiques.
Gènes - groupe ethnique – prédisposition – changement d’environnement - origine génétique marquée – jumeau - 90 %
Le mode de vie de l’individu influence-t-il aussi la survenue de ce diabète ?
B. Facteurs environnementaux.
Obésité - sédentarité - diabète gras - milieu urbain – hypercalorique - milieu rural - hypocalorique
Les diabètes résultent de l’interaction entre ces gènes et des facteurs de l’environnement, en particulier l’alimentation.
Connaissant les principales causes de ce diabète, comment peut-on prévenir la maladie ?
II. La prévention du diabète de type 2.
Pour lutter efficacement contre les diabètes il est nécessaire :
- d'identifier des sujets à haut risque ;
- d'essayer de prévenir l'apparition de la maladie ;
- et enfin de traiter la maladie si elle apparaît.
Ici ce sont les deux premiers points qui nous intéressent.
A. Identifier les sujets à haut risque.
Gènes de prédisposition, de susceptibilité études familiales polymorphisme médecine prédictive.
Quelles précautions doit prendre un individu à risque pour éviter de développer la maladie ?
B. Prévenir l'apparition de la maladie.
Régime alimentaire - modifier la masse graisseuse - sensibilité des adipocytes à l'insuline.
Activité physique lutte contre l'insulinorésistance au niveau musculaire.
Conclusion : multifactoriel prévention et problèmes éthiques.
Si la prévention a échoué, comment peut-on aider le diabétique dans sa vie quotidienne ? |
| la régulation de la glycémie et les phénotypes diabétiques |
Sujet 3
Introduction :
- Amorce : 2 types de diabète, régulation défaillante
- Pb : Comment peut-on expliquer le phénotype diabétique de type 2 ?
- Annonce du plan : régulation = schéma ; phénotype diabétique de type 2 ; origines.
I. La régulation de la glycémie : schéma bilan.
Système réglé |
- Milieu intérieur : homéostasie
- Paramètre réglé = glycémie
|
|
perturbation |
Repas : hyperglycémie
Jeûne : hypoglycémie |
Système réglant |
Capteur = ilot de Langerhans |
- Cellules ß centrales
- Cellules a périphériques
|
Système de transmission = hormones |
|
Organes effecteurs |
cellules hépatiques :
• glycogénogenèse + synthèse AG
• ou glycogénolyse et néoglucogenèse
cellules musculaires : glycogénogenèse
adipocytes : synthèse d'AG (puis triglycérides) |
boucle |
de régulation : |
retour à la valeur de consigne |
récepteurs |
spécifiques |
|
II. Le phénotype diabétique de type 2.
- Diabète de l'âge mûr ou diabète gras.
- L'hyperglycémie.
- Le DNID (récepteurs, insulinorésistance, déficit en insuline secondaire)
III. Origines de ce phénotype.
- Facteurs environnementaux. (obésité ; sédentarité)
- Facteurs génétiques. (très marqués : jumeaux ; gènes de prédisposition ; mono ou polygénique)
Conclusion : résumé : diabète dû mauvaise régulation. Elargissement : diabète de type 1 a mêmes causes ? |
|
|
La part du génotype et la part de l'expérience individuelle dans le fonctionnement du système nerveux.
|
Corrigé |
|
Un réflexe myotatique, comme les réflexes achilléen et rotulien, est déclenché par l’étirement d’un muscle extenseur, lors d’une position en déséquilibre, par exemple. Cette stimulation provoque la contraction involontaire de ce même muscle et le relâchement de son antagoniste. Il en résulte un retour à l’équilibre et le maintien de la posture.
Voir schéma du cours ou p 392 du livre.
|
|
La genèse d’un potentiel d’action repose sur l’existence d’un potentiel de repos : l’extérieur de la membrane est chargé positivement par rapport à l’intérieur.
Suite à une stimulation, un message nerveux peut naître :
- si l’intensité de la stimulation est faible, inférieure au seuil de stimulation, elle produit une faible dépolarisation, de quelques mV, au niveau de la fibre nerveuse .
- lorsque l’intensité de la stimulation augmente, l’amplitude de cette dépolarisation devient brusquement (quelques ms) très importante, au point de provoquer une inversion du potentiel de membrane (jusqu’à +20 à + 35 mV selon le type de fibre) : c’est le potentiel d’action ;
- si l’intensité de la stimulation est encore plus forte, ce n’est pas 1 mais 2, 3 ou plus de potentiels d’actions (train de PA) qui sont créés.
Ces PA ont tous la même amplitude : la fibre répond à la loi du « tout ou rien ». Le message nerveux est codé en fréquence des PA.
|
|
Introduction :
Le message nerveux est un message codé qui renseigne l’organisme sur, notamment, des informations de l’environnement ou sur des conditions régnant dans l’organisme. Par exemple, il renseigne nos centres nerveux sur l’état d’étirement de certains muscles de manière à ce que ces centres nerveux commandent les contractions appropriées pour le maintien de la posture dans le cadre du réflexe myotatique.
Le stimulus et son intensité sont codés au niveau d’un récepteur sensoriel et le message afférent ainsi créé se propage le long d’une fibre nerveuse sensitive puis est transmis à un motoneurone au niveau d’une synapse ; on l’appelle alors message efférent.
Schéma du trajet du message nerveux lors d’un arc réflexe myotatique
(= réponse d’un muscle à son propre étirement)

I Codage de l’intensité du stimulus au niveau du récepteur : le potentiel de récepteur
A. Le potentiel de repos
Une microélectrode enregistre un potentiel de 0 mV en surface et –70 mV à l’intérieur de l’axone. Cette différence de potentiel de 70 mV de part et d’autre de la membrane de l’axone est permanente : on l’appelle potentiel de repos. L’intérieur de la cellule est électronégatif par rapport à l’extérieur : la membrane est polarisée.
Cette polarisation ou potentiel de repos est due à une dissymétrie ionique maintenue par la membrane. Ces potentiels membranaires peuvent être enregistrés pour tout type de cellule.
Exemple :
- neurone –70 mV
- cellule musculaire –90 mV

L'enregistrement obtenu quand une microélectrode est plongée dans l’axoplasme (entre t1 et t2) par rapport à un électrode de référence dans le milieu extracellulaire :

B. Naissance du potentiel de récepteur
Le récepteur, ici le fuseau neuromusculaire, constitué de fibres musculaires modifiées entourées par des terminaisons nerveuses sensitives, réagit à un stimulus déterminé, mécanique, l’étirement d’un muscle.
Le stimulus déclenche une modification de l’état électrique du neurone sensitif. Cette dépolarisation ou potentiel de récepteur est graduable est fonction de l’intensité du stimulus et est localisée (non propageable).
intensité de stimulation i1< i2 < i3
La polarisation de repos de la membrane tend à s’inverser.
Si ce PR dépasse le seuil de dépolarisation de la fibre, il en résulte la genèse d’un PA.
II Le message propagé le long du neurone sensitif : les potentiels d’action
Lors d’une stimulation de faible intensité (i1, i2), pas de PA. La dépolarisation de la membrane de la fibre nerveuse augmente avec l’intensité de la stimulation.
A partir de i3, PA : c’est le seuil de dépolarisation nécessaire pour faire naître un PA
i4>i3 mais le PA garde la même amplitude : c’est la loi du tout ou rien.
Les neurones n’émettent un PA que si leur membrane est dépolarisée jusqu’à une valeur seuil : le seuil de dépolarisation. Au delà de ce seuil, l’amplitude des PA émis reste constante : c’est la loi du tout ou rien.
Potentiel d’action d’une fibre nerveuse en réponse à une stimulation isolée

Un PA est un signal élémentaire du message nerveux. Il correspond à une modification du potentiel de repos. (amplitude de l’ordre de 100 mV ; durée : 1 à 2 ms). Il comprend :
- une phase ascendante au cours de laquelle le potentiel transmembranaire passe d’une valeur négative (potentiel de repos) à une valeur positive = dépolarisation
- une phase descendante au cours de la quelle le potentiel passe d’une valeur positive à des valeurs de plus en plus négatives jusqu’à atteindre le potentiel de repos = repolarisation puis hyperpolarisation.
Les PA enregistrés sont tous de même amplitude quelle que soit l’intensité de la stimulation ; c’est la fréquence des PA qui augmente avec l’intensité de stimulation. Au niveau d’un neurone, le message nerveux se traduit par une succession de potentiels d’action d’amplitude constante et dont la fréquence code l’intensité de la stimulation.
III Transmission synaptique au motoneurone
Le message est transmis d’un neurone à l’autre grâce à une substance chimique : un neurotransmetteur libéré par le neurone présynaptique par exocytose dans des concentrations qui traduisent la fréquence des PA qui parvient à la terminaison de ce neurone. La fixation des molécules du NT aux récepteurs de la membrane postsynaptique induit un changement du potentiel membranaire nommé potentiel postsynaptique dont l’amplitude varie en fonction de la quantité de NT reçue et qui déclenche la création d’un PA au niveau d’un neurone quand le seuil de dépolarisation de celui-ci est atteint.
L’action des molécules du NT est très fugace (il est rapidement éliminé de l’espace synaptique), ce qui autorise une modulation très fine de l’activité de la cellule postsynaptique.
Au niveau d’une synapse, le message codé en fréquence est donc traduit en un message chimique codé en concentration de NT, lui-même codé en amplitude au niveau postsynaptique sous forme d’un PPS qui déclenchera, à partir d’une amplitude seuil, un PA qui sera la base d’un message codé en fréquence et propagé le long du motoneurone.
Schéma du fonctionnement synaptique.

Conclusion :
Le message nerveux est d’abord de nature électrique, un potentiel codé en amplitude (potentiel de récepteur) puis en fréquence le long de la fibre sensitive (PA). Au niveau d’une synapse, la transmission se fait sous forme d’un message chimique (neurotransmetteur) qui permet de créer un nouveau message électrique codé en fréquence le long du motoneurone.
|
|
Intro : définition du message nerveux (train de PA). Schéma de l’arc réflexe.
I. Intervention de la membrane plasmique dans la naissance du message nerveux.
- Le potentiel de repos : une polarisation membranaire en absence de stimulation.
- Le potentiel d’action : une dépolarisation brutale et fugace.
Loi du tout ou rien – Seuil de dépolarisation.
II. La membrane plasmique et la conduction du message nerveux.
- La propagation au niveau des fibres amyéliniques.
- La propagation au niveau des fibres myélinisées.
III. La membrane plasmique et la transmission du message.
- Les membranes pré et postsynaptiques.
- Le fonctionnement synaptique.
Conclusion : rôle essentiel des membranes neuronales. Elargissement. |
la transmission synaptique et l'intégration |
Intro : propagation des messages nerveux le long des fibres nerveuses jusqu’aux extrémités synaptiques.
I. Le fonctionnement de la synapse neuroneuronique.
A. Transmission synaptique : explication + schéma.
Message chimique – action rapide et fugace (dégradation des neurotransmetteurs)
B. Le codage du message : concentration en neurotransmetteurs.
C. Les différents types de synapse : excitatrice (dépolarisation), inhibitrice (hyperpolarisation).
II. Originalité du message postsynaptique.
A. Sommation spatiale.
B. Sommation temporelle.
C. Intégration et naissance ou pas d’un message postsynaptique.
|
la transmission synaptique et l'intégration |
Introduction : Les neurones sont des cellules spécialisées dans la propagation et la transmission des messages nerveux. C’est au niveau des synapses qu’un message nerveux est transmis d’un neurone à l’autre.Les neurones connectés sont le plus souvent séparés par un espace (fente synaptique), ce qui pose le problème de la transmission des potentiels d’action (P.A.) qui constituent le message nerveux. - Nous verrons donc dans un premier temps la transmission synaptique et dans un deuxième temps l’intégration par le neurone postsynaptique.
I. La transmission synaptique.
A. Fonctionnement d’une synapse chimique.
- Schéma.
- Expliquer les différentes étapes : arrivée des P.A., libération N.T. (codage), fixation du N.T. sur les récepteurs spécifiques, création P.P.S., destruction du N.T.
B. La réponse du neurone postsynaptique.
Schéma d’un P.P.S.E. et d’un P.P.S.I. et explication.
II. L’intégration par le neurone postsynaptique.
A. La sommation spatiale : schéma + explication.
B. La sommation temporelle.
C. Naissance d’un P.A. propagé.
Conclusion : Les synapses constituent une discontinuité empêchant le franchissement d’un message nerveux électrique, elles fonctionnent donc de façon chimique. Cela implique l’unidirectionnalité de la circulation du message nerveux et le traitement de l’information. Le cerveau, avec ses milliards de neurones, constitue une machinerie très complexe, dont le fonctionnement est encore loin d’être élucidé..
|
la plasticité cérébrale |
|
|
Comparaison système nerveux et hormonal (pour avoir une idée plus globale des 2 parties précédentes : pas facile !)
Corrigé |
Intro : La communication entre les cellules d’un organisme pluricellulaire est nécessaire pour régler leur développement et leur organisation en tissus, pour contrôler leur croissance et leur division et pour coordonner leurs fonctions.
La transmission de l’information met en jeu deux types des messages :
- les uns nerveux, générés au niveau des fuseaux neuromusculaires dans le cas du réflexe myotatique ;
- les autres hormonaux, générés au niveau de cellules endocrines comme celles du pancréas dans le cas de la régulation de la glycémie.
Dans ce devoir, nous étudierons dans un premier temps les facteurs qui déclenchent l’activité des neurones et des cellules endocrines avant de nous attarder sur les caractéristiques des messages émis.
I. Les facteurs déclenchant l’activité des neurones et des cellules endocrines.
A. La stimulation du fuseau neuromusculaire.
La percussion du tendon d’Achille, par exemple, provoque l’étirement des fibres constituant le muscle stimulé*. Certaines de ces fibres musculaires sont entourées de terminaisons dendritiques de neurones sensitifs, dont le corps cellulaire se situe dans la racine dorsale du nerf rachidien et la terminaison synaptique au niveau de la substance grise de la moelle épinière au sein de laquelle il fait synapse avec un neurone moteur. L’étirement provoque la naissance d’un message nerveux sensitif dans ces neurones, train de potentiels d’action
B. La stimulation d’une cellule endocrine.
Le pancréas est sensible aux variations de la glycémie par rapport à une grandeur consigne voisine de 1g.L -1. En effet, il possède des cellules capables de détecter les écarts par rapport à cette valeur de référence. Après stimulation, la cellule endocrine voit sa synthèse protéique stimulée, l’amenant à produire puis excréter des hormones par exocytose.*
La stimulation de ces deux types cellulaires repose sur des mécanismes différents mais a la même conséquence : leur fonctionnement est modifié. Mais le message émis alors sera-t-il différent ?
II. Les caractéristiques du message émis.
A. Nature et codage.
- Nature et codage du message nerveux.
Nous avons vu précédemment que le message nerveux est de nature électrique. Il s’agit en effet de séries de potentiels d’action* c'est-à-dire de dépolarisations brutales suivies de repolarisations rapides, et enfin hyperpolarisations. Le codage est réalisé en fréquence des potentiels d’action : plus l’intensité de la stimulation est forte, plus le message comporte de nombreux PA dans un temps donné.
Toutefois, au niveau de la zone de contact entre deux neurones successifs (ou entre un neurone et un muscle), il n’y a pas continuité et ceci implique une communication chimique grâce à des neurotransmetteurs (nous détaillerons cette communication en C1). Le codage est ici réalisé en concentration de neurotransmetteur.
- Nature et codage du message hormonal.
Le message hormonal est de nature chimique, il s’agit de molécules comme l’insuline ou le glucagon dans le cas de la régulation de la glycémie. Le codage est ici réalisé en concentration hormonale c'est-à-dire suivant la quantité d’hormone par volume de sang (à noter que les quantités sont toujours extrêmement faibles).
B. Propagation.
- Propagation du message nerveux.
La propagation se réalise sans atténuation car les dépolarisations se recréent le long de la membrane de la fibre nerveuse qui oriente la circulation du message. La vitesse est augmentée par la présence d’une gaine de myéline* entourant la fibre nerveuse : de 10 à 100 m.s -1.
- Propagation du message hormonal.
Les hormones sortent de la cellule productrice par exocytose et sont déversées dans un vaisseau sanguin. Elles parcourent alors tout l’organisme par le biais de la circulation sanguine. Cette circulation est assez lente, quelques minutes pour des organes voisins.
C. Transmission aux cellules effectrices.
- Le fonctionnement synaptique.*
Les motoneurones transmettent leur message aux fibres musculaires du muscle stimulé et un message opposé au muscle antagoniste. Cette fois encore intervient une communication chimique. En effet l’arrivée du message nerveux provoque l’exocytose des vésicules de neurotransmetteurs (d’autant plus de vésicules que le message était à haute fréquence), molécules qui se fixent sur leurs récepteurs spécifiques au niveau de la membrane postsynaptique. Cette fixation déclenche l’entrée d’ions positifs ou négatifs. S’il s’agit d’ions positifs en forte quantité, un nouveau message électrique est généré et le muscle se contracte.
- Les cellules cibles des hormones.
Les hormones libérées dans le sang vont agir sur les seules cellules qui possèdent des récepteurs de configuration spatiale complémentaire à l’hormone* (cellules hépatiques mais aussi adipeuses et musculaires pour l’insuline). Cela modifie leur fonctionnement, dans l’exemple présenté il y a stockage de glucose sous forme de glycogène ou acides gras.
Conclusion : Malgré des mécanismes différents dans les deux modes de communication ici envisagés, nous avons constaté certaines ressemblances apparaissant dans le tableau ci-dessous :
Mode de communication |
Nerveuse |
Hormonale |
Facteurs déclenchant |
Stimulus = étirement du fuseau neuromusculaire |
Ecart par rapport à la valeur consigne (perturbation de l’homéostasie) |
Nature des messages |
Electrique, chimique (synapse) |
chimique |
Codage des messages |
Fréquence des PA
Concentration en neurotransmetteurs |
Concentration hormonale |
Propagation |
Rapide, le long des neurones |
Lente, via la voie sanguine |
Transmission |
Molécules de neurotransmetteurs et récepteurs spécifiques |
Hormones et récepteurs spécifiques des cellules cibles |
* schéma du cours |
Intro : La communication entre les cellules d’un organisme pluricellulaire est nécessaire pour régler leur développement et leur organisation en tissus, pour contrôler leur croissance et leur division et pour coordonner leurs fonctions.
La transmission de l’information met en jeu deux types des messages :
- les uns nerveux, générés au niveau des fuseaux neuromusculaires dans le cas du réflexe myotatique ;
- les autres hormonaux, générés au niveau de cellules endocrines comme celles du pancréas dans le cas de la régulation de la glycémie.
Quels sont les points communs et différences entre ces deux types de communication ?
Dans ce devoir, nous comparerons ces deux types de messages sous les aspects suivants : voies de communication puis nature des signaux, codage de l’information, et enfin performances.
I. Les voies de communication.
A. Pour le message nerveux.
Le message nerveux est généré au niveau de récepteurs sensoriels comme les fuseaux neuromusculaires* et emprunte les nerfs rachidiens reliés à la moelle épinière. Ces nerfs sensitifs sont constitués de neurones en T : long dendrite du récepteur au ganglion rachidien, corps cellulaire dans le ganglion et axone se terminant dans la substance grise de la moelle épinière. A ce niveau, ils font synapse d’une part avec des motoneurones qui aboutissent au muscle stimulé au niveau de plaques motrices (ou synapses neuromusculaires), d’autre part avec des interneurones inhibiteurs qui sont en relations avec des motoneurones aboutissant au muscle antagoniste.
B. Pour le message hormonal.
Les hormones sont véhiculées par voie sanguine et parcourent donc tout l’organisme dans le système circulatoire.
La communication nerveuse est donc guidée puisque le parcours est imposé par les circuits neuronaux, par contre la communication hormonale, se réalise à distance.
Mais quel type de message est propagé ?
II. La nature des signaux.
A. Pour le message nerveux.
Le message nerveux est de nature électrique. Il s’agit en effet de séries de potentiels d’action c'est-à-dire de dépolarisations brutales suivies de repolarisations rapides, et enfin hyperpolarisations. *
Toutefois, au niveau de la zone de contact entre deux neurones successifs (ou entre un neurone et un muscle), il n’y a pas continuité et ceci implique une communication chimique grâce à des neurotransmetteurs.
Au niveau d’une synapse*, l’arrivée du message nerveux provoque l’exocytose des vésicules de neurotransmetteurs, molécules qui se fixent sur leurs récepteurs spécifiques au niveau de la membrane postsynaptique. Cette fixation déclenche l’entrée d’ions positifs ou négatifs. S’il s’agit d’ions positifs en forte quantité, un nouveau message électrique est généré (et le muscle se contracte s’il s’agit d’une plaque motrice).
B. Pour le message hormonal.
Le message hormonal est de nature chimique, il s’agit de molécules comme l’insuline ou le glucagon dans le cas de la régulation de la glycémie.
Une différence nette dans la nature des signaux est donc à noter mais la communication synaptique, chimique, présente des points communs avec la communication hormonale.
Mais comment est codé le message dans les deux cas ?
III. Le codage de l’information.
A. Pour le message nerveux.
Le codage est réalisé en fréquence des potentiels d’action : plus l’intensité de la stimulation est forte, plus le message comporte de nombreux PA dans un temps donné.
Toutefois, au niveau de la zone de contact entre deux neurones le codage est réalisé en concentration de neurotransmetteur : il y a d’autant plus de vésicules qui libèrent leur contenu que le message électrique était à haute fréquence.
B. Pour le message hormonal.
Le codage est ici réalisé en concentration hormonale c'est-à-dire suivant la quantité d’hormone par volume de sang (à noter que les quantités sont toujours extrêmement faibles).
Un codage en fréquence s’oppose à un codage en amplitude, toutefois on retrouve au niveau synaptique un phénomène comparable à la communication hormonale.
Mais les performances de ces communications sont-elles comparables ?
IV. Les performances.
A. Pour le message nerveux.
Le message nerveux, de par sa nature électrique, est extrêmement rapide : de l’ordre de 10 m.s -1 pour un petit axone « nu » contre 100 m.s -1 pour une fibre myélinisée. Toutefois cette rapidité est diminuée par la transmission synaptique qui freine le message, de manière modérée toutefois : 0,5 millisecondes sont nécessaires au franchissement de la synapse, on parle de délai synaptique.
C’est un message qui est fugace c'est-à-dire qui ne dure pas, la fixation des neurotransmetteurs sur leurs récepteurs étant notamment très brève.
B. Pour le message hormonal.
Dans ce cas, on ne parle plus en millisecondes mais en minutes voir en heures. Cela est dû à la circulation « aléatoire » des hormones qui parcourent l’ensemble du circuit sanguin et, « parfois », se fixent aux récepteurs spécifiques que possèdent certaines cellules appelées cellules cibles.
C’est un message plus durable que le message nerveux.
Conclusion : Ces deux modes de communication se complètent dans l’organisme et interviennent même parfois ensemble dans certaines régulations (pression artérielle par exemple). Ils présentent un certain nombre de points communs et de différences répertoriés dans le tableau suivant :
Mode de communication |
Nerveuse |
Hormonale |
Voies de communication |
Neurones |
sang |
Nature des signaux |
Electrique, chimique (synapse) |
chimique |
Codage de l’information |
Fréquence des PA
Concentration en neurotransmetteurs |
Concentration hormonale |
Performances |
Rapide, fugace |
Lente, durable |
* schéma du cours |
| Intro : Les deux modes de communication, nerveux et hormonal, se complètent dans l’organisme et interviennent même parfois ensemble dans certaines régulations (pression artérielle par exemple). Ils présentent un certain nombre de différences mais aussi de points communs en particulier au niveau de la transmission chimique du message.
Nous examinerons dans un premier temps les différences dues aux différences de distance avant de rechercher les ressemblances au travers de la comparaison de la transmission endocrine avec la transmission synaptique.
I. Différences : mécanismes et vitesse.
A. Dans la transmission endocrine.
Les cellules endocrines spécialisées, comme les cellules du pancréas dans la régulation de la glycémie, sécrètent, suite à la détection d’un écart par rapport à la valeur consigne de la variable à régler (ici la glycémie), des hormones qui circulent dans le sang pour agir sur des cellules cibles dispersées dans l’organisme. Il y a action à distance, la vitesse est donc peu importante, il faut quelques minutes pour qu’une hormone atteigne sa cellule cible.
B. Dans la transmission synaptique.
Les extrémités de neurones, dont la membrane est parcourue par une série de dépolarisations ou potentiels d’action, libèrent des neurotransmetteurs au niveau de structures spécialisées appelées synapses chimiques. Les neurotransmetteurs diffusent à travers la fente synaptique, dont la largeur est d’environ 50 nm et n’agit que sur la cellule cible postsynaptique adjacente. C’est une communication de proximité, extrêmement rapide, de l’ordre d’une ms pour franchir une synapse.
Ces deux stratégies de communication intercellulaire sont différentes selon les distances auxquelles ils agissent. Peut-on toutefois trouver des ressemblances ?
II. Ressemblances : la réponse de la cellule cible.
A. Dans la transmission endocrine.*
La spécificité de la transmission dans le système endocrinien dépend des propriétés chimiques du signal (hormone) et des récepteurs de la cellule cible : chaque type de cellule endocrine sécrète une hormone différente dans le sang et chaque cellule qui porte des récepteurs complémentaires va répondre de façon qui correspond au type cellulaire. Par exemple, l’insuline n’agira que sur les cellules possédant des récepteurs de conformation spatiale complémentaire, à savoir les cellules hépatiques, adipeuses et musculaires. Ces cellules voient alors leur fonctionnement modifié, elles stockent le glucose sous forme de glycogène ou d’acides gras. Par contre le glucagon ne peut se fixer qu’aux cellules hépatiques, seules à posséder les récepteurs appropriés. Il a pour conséquence la libération de glucose à partir des réserves de glycogène du foie.
B. Dans la transmission synaptique.*
Les cellules nerveuses peuvent transmettre l’information à longue distance le long des fibres nerveuses. Les neurotransmetteurs, libérés dans la fente synaptique, se fixent sur leurs récepteurs spécifiques, comme dans le cas de la communication hormonale. Cette fixation provoque, elle aussi, une modification de la cellule postsynaptique, naissance d’une nouvelle dépolarisation ou hyperpolarisation c'est-à-dire un changement de la polarité membranaire. Cette dernière peut donner naissance à un nouveau message nerveux postsynaptique ou une contraction musculaire.
Conclusion : Les transmissions chimiques endocrines et synaptiques présentent donc de nombreux points communs. Toutefois on peut noter que les cellules nerveuses fonctionnent avec beaucoup plus de précision, l’information étant plus « guidée » alors que les hormones sont fortement diluées dans le flux sanguin et doivent cependant être capables d’agir à de très faibles concentrations.
* schéma du cours |
|
|
| SCIENCES DE LA TERRE : Structure, composition et dynamique de la Terre. |
Structure et composition chimique de la Terre interne.
Corrigé |
On peut connaître la composition de "la Terre accessible" en observant les paysages, des affleurements, en ramassant des échantillons de roches.
Comment la Terre est-elle constituée ?
I. Un modèle de la Terre accessible : étude de la Terre en surface.
A. Au niveau d’un paysage.
Quelles informations relatives à la structure interne de la Terre l'étude des paysages peut-elle nous apporter ?
L’analyse d’un paysage permet :
- de révéler quelques grandes structures géologiques (volcans, reliefs, accidents tectoniques : failles) qui témoignent d’une Terre active;
- de mettre en relation des données d’observation avec des données cartographiques.
- de chercher un endroit où les matériaux du sous-sol sont accessibles à l’observation et à l’échantillonnage : un affleurement : par exemple une falaise sur le littoral, une carrière creusée par l’homme, ou encore une tranchée sur le bord de la route…
Quelles informations peut nous apporter l’étude d’un affleurement ?
B. Au niveau d’un affleurement.
L’étude d’un affleurement commence toujours par un croquis sur lequel on distingue le sol du sous-sol, on repère l’organisation géométrique des roches qui affleurent, leurs couleurs, les strates en relief, la fracturation
Le deuxième temps peut être consacré à la description de chaque unité identifiée :
- l’aspect général des roches de même nature (massive, feuilletée…), leur réaction à l’acide par exemple ;
- caractéristiques géométriques (inclinaison, cassures, plissements…), couleurs, …
- observation d’indices interprétables grâce au principe de l’actualisme : des fossiles, fentes de dessication, stromatolites…
Actualisme (ou uniformitarisme) : théorie postulant que les lois régissant les phénomènes géologiques actuels étaient également valables dans le passé (principe des causes actuelles et des causes anciennes).
Une fois ces éléments notés, des échantillons représentatifs de chaque unité repérée peuvent être prélevés pour une étude ultérieure au laboratoire et à l’échelle microscopique.
NB : Il faut impérativement respecter le site et les matériaux étudiés. (A revoir lors de la sortie sur le terrain)
L’étude d’un affleurement à la surface de la Terre permet d’observer les matériaux de la croûte et parfois du manteau supérieur.
Que nous apporte l’étude des roches prélevées au niveau des affleurements ?
C. Au niveau des roches.
1) Les roches de la croûte continentale.
L’étude d’une carte de répartition des grands types de roches montre que les continents sont constitués de trois grands types de roches :
- les roches sédimentaires (argiles, calcaires, grès…), disposées en couches superposées = strates car formées sous l’eau.
- les roches magmatiques : le granite, le basalte.
- les roches métamorphiques : le gneiss.
Nous allons étudier un exemple de roche de la croûte continentale : le granite.
a. Etude de l’échantillon de granite.
C’est une roche grenue c'est-à-dire formée de minéraux jointifs, visibles à l’œil nu :
- du quartz : blanc sale, xénomorphe
- du micas noir = biotite : noir brillant
- des feldspaths :
- orthose : rose
- plagioclases : blanc, automorphe.
- Etude de la lame mince.
Le granite est issu d’un refroidissement lent consécutif à une montée à vitesse régulière du magma vers la surface de la croûte continentale.
b. La composition chimique du granite.
Les roches de la croûte continentale sont constituées de minéraux silicatés riches en silice et en Al, Na, K et Ca.
c. La densité du granite.
La densité = quotient de la masse volumique d’un corps sur la masse du même volume d’un fluide de référence (eau en général).
La masse volumique = quotient de la masse d’un corps sur son volume.
On peut déterminer la masse volumique d’un échantillon en le plongeant dans l’eau et en mesurant l’augmentation du volume d’eau.
Masse volumique du granite 2,5 g/cm 3
2) Les roches des fonds océaniques.
Sous un plancher de sédiments, les fonds océaniques sont constitués de trois grands types de roches, de haut en bas :
- des basaltes en coussins ;
- des gabbros ;
- des péridotites.
a. Le basalte.
L’observation d’une lame mince de basalte montre que cette roche, de structure (ou texture) microlitique, est constituée de verre (non cristallisé) et de microlithes de pyroxène, d’olivine et de feldspaths plagioclases.
Le basalte est donc issu d’un refroidissement brutal (trempe), consécutif à une montée du magma vers la croûte océanique. La forme en coussins prouve cette trempe au contact de l’eau froide.
Le basalte est également constitué de silicates riches en Ca et Mg.
La masse volumique du basalte est proche de celle du granite : 2,5 g/cm 3
b. Le gabbro.
La composition chimique est la même que celle du basalte, cela prouve l’origine commune, les deux roches sont formées à partir du même magma.
Toutefois la structure est différente, le gabbro est une roche grenue car elle a cristallisé en profondeur.
c. La péridotite : une roche du manteau.
Localement de la péridotite affleure, sous les gabbros ou directement au fond de l’océan au niveau de failles.
Le manteau supérieur est constitué de péridotite, roche grenue constituée principalement de deux minéraux : l’olivine et le pyroxène (+ amphibole, biotite, grenat)
Les minéraux de la péridotite du manteau supérieur sont des silicates pauvres en silice mais riches en fer et en magnésium.
Masse volumique de la péridotite 3,5 g/cm3
Conclusion :
Les enveloppes de la Terre, accessibles par échantillonnage, ont des compositions chimiques différentes que l’on détermine à partir de l’étude des roches représentatives.
Ces roches représentatives sont formées de minéraux et/ou de verre. Les principaux minéraux sont : olivines, pyroxènes, feldspaths, quartz, amphiboles et micas.
La composition chimique des roches est présentée en % massique d’éléments chimiques.
La composition chimique des enveloppes de la Terre est dominée par un nombre limité d’éléments dits « majeurs » (Si, O, Mg, Fe, Ca, Na, K, Al). Ces éléments sont hébergés dans les principaux minéraux des roches. |
Pb : Comment les météorites nous renseignent sur la structure du globe ?
I. Trois grands groupes de météorites.
- les chondrites (85 % des météorites) sont riches en silicates et en fer ;
- les achondrites sont riches en olivine et pyroxène.
- les sidérites, ou achondrites métalliques, très denses, sont riches en fer et nickel.
II. Les informations apportées par les météorites.
1) Formation des objets du système solaire.
La Terre , comme les autres planètes du système solaire, s’est formée il y a 4,6 milliards d’années par accrétion, à partir d’une nébuleuse.
Toutefois, des objets rocheux ne se sont pas agglomérés, il s’agit des astéroïdes.
Les astéroïdes qui quittent leur orbites et « s’échouent sur Terre constituent donc des vestiges des roches constitutives de la Terre, accessibles à l’observation et à l’analyse chimique.
Comment se sont formées ces météorites ?
2) Origine des météorites.
Une météorite s’est formée par accrétion de gaz et de poussières qui étaient présents dans la nébuleuse ayant donné naissance à notre système solaire, exactement comme la Terre. Certaines fondent et se différencient avant d’être fragmentées (achondrites).
Par analogie, on admet aujourd’hui que la terre, de – 4,6 à – 4,4 Ma, s’est différenciée avec :
- fusion d’un volume important de la Terre sous l’action de la chaleur interne et des impacts météoritiques très nombreux durant cette période ;
- le fer et le nickel, plus denses, migrent vers l’intérieur de la Terre (formation du noyau) comme en attestent les sidérites ;
- les silicates fondus cristallisent et édifient le manteau (comme en attestent les achondrites) ;
- une atmosphère se forme par dégazage.
Conclusion : Les matériaux du manteau profond et du noyau sont inaccessibles. On peut néanmoins, par des modèles et des raisonnements qui tiennent compte de la formation de la Terre à partir des chondrites, préciser leurs compositions.
La structure de la Terre, résulte d’une part de sa formation par accrétion de petits corps dont les météorites de type chondrite sont les vestiges, d’autre part de sa différenciation.
|
Pour voir le fœtus dans le ventre de sa mère, on utilise l’échographie, c'est-à-dire que l’on envoie des ultrasons, réfléchis par les organes du futur bébé. Pour la terre, on étudie les séismes naturels ou on provoque des vibrations du sol.
Comment peut-on connaître la Terre en profondeur ?
I. Séismes et ondes sismiques.
(à la limite du sujet)
Un séisme ou tremblement de terre se traduit en surface par des vibrations du sol. Il provient de la fracturation des roches en profondeur. Cette fracturation est due à une grande accumulation d’énergie qui se libère, en créant ou en faisant rejouer des failles, au moment où le seuil de rupture mécanique des roches est atteint.
On détecte les vibrations du sol, appelées ondes sismiques, grâce à des sismomètres qui enregistrent des sismogrammes.
Un sismogramme est composé 3 trains d’ondes successifs :
- les ondes P (ou premières) de compression et décompression ( grondement lors d’un séisme).
- les ondes S (secondes) de cisaillement (ou transversales).
- les ondes L ne se propagent qu’en surface.
Puisque nous voulons mieux comprendre comment nous avons trouvé la structure profonde de la Terre, nous n’allons utiliser que les ondes qui se propagent en profondeur (P et S).
Quelles informations nous apporte l'étude des ondes P et S sur la structure profonde du globe ?
II. Propriétés des ondes sismiques et structure du globe.
1) Un constat : une zone d’ombre.
Quel que soit le lieu du séisme, tous les sismographes situés dans une bande entre 104 et 144° (angle à l’épicentre) ne reçoivent aucune onde directe : on parle de zone d’ombre.

Comment peut-on expliquer cette disparition des ondes P (zone d’ombre) ?
2) Modélisation du trajet des ondes sismiques.
La propagation d’une onde sismique est comparable à celle d’un rayon lumineux on peut donc construire un modèle analogique pour comprendre ce qui se passe à l’intérieur du globe.
Le bord du petit cristallisoir représente une surface de discontinuité.
Lorsqu’un « rai sismique » (analogie avec un rayon lumineux) atteint la surface entre deux milieux de natures différentes (interface), sa trajectoire est déviée : il est soit réfléchi (ne traverse pas), soit réfracté (passe mais avec une déviation de la trajectoire), soit les deux.
La surface entre deux milieux de propriétés différentes constitue une discontinuité.
L’étude de la propagation des ondes sismiques (en appliquant la loi optique de Descartes) permet de calculer, connaissant le temps d’arrivée des ondes, les vitesses de propagation et la profondeur de la discontinuité.
3) Vitesse des ondes sismiques et structure du globe.
On observe des trajectoires courbes qui s’expliquent par des déviations des rais sismiques mais toujours pas de zone d’ombre.
L’existence de la zone d’ombre repose donc sur la présence d’une discontinuité située à 2900 km de profondeur, qui sépare le noyau central, du manteau périphérique.
Le graphe de variation des vitesses des ondes en fonction de la profondeur construit par les géologues est complexe, il fait apparaître plusieurs discontinuités (brutales variations de vitesse) :
- la discontinuité de Mohorovicic appelée couramment le Moho à quelques dizaines de km de profondeur [vers 5 km (sous les fonds océaniques) à 70 km (sur les continents)] au-delà de laquelle les ondes s’accélèrent, il s’agit de limite entre la croûte et le manteau .
- une discontinuité vers 100- 120 km au-delà de laquelle les ondes ralentissent, c’est la limite entre la lithosphère et l’asthénosphère .
- une discontinuité « légère » vers 700 km de profondeur, c’est la limite entre l’asthénosphère et le manteau inférieur (zone de nouveau très rigide).
- la discontinuité de Gutenberg marquant la limite entre manteau inférieur et noyau externe , caractérisée par l’absence des ondes S ( 2 900 km).
- la discontinuité de Lehman à 5 150 km de profondeur, entre noyau externe et le noyau interne solide (les ondes P ont une vitesse constante, les ondes S « réapparaissent »), on l’appelle la graine.
L’étude de la propagation des ondes sismiques montre que la Terre est structurée en enveloppes concentriques de tailles, masses et masses volumiques différentes : la croûte (continentale ou océanique), le manteau et le noyau séparés par des discontinuités.
A quels changements physico-chimiques correspondent ces discontinuités ?
III. Interprétation des données sismiques.
Nous savons déjà que :
- La discontinuité de Moho correspond à un changement de composition chimique de matériaux. (basalte / gabbro ou granite / gneiss pour la croûte et péridotite pour le manteau)
- La discontinuité de Gutenberg est une discontinuité physique entre le manteau solide riche en péridotite et le noyau externe liquide puisque les ondes S ne la traversent pas.
- La discontinuité de Lehman est une discontinuité physique entre le noyau externe liquide (fer et Nickel) et le noyau interne solide plus dense.
Les études de laboratoire, en reconstituant les conditions subies par les matériaux, permettent d’interpréter les données sismiques pour déduire d’autres propriétés du globe :
- la pression qui règne en profondeur ;
- la densité des roches en profondeur ;
- la composition chimique des couches profondes.
1) Informations complémentaires sur la discontinuité de Gutenberg.
La discontinuité de Gutenberg est aussi une discontinuité chimique entre le manteau riche en péridotite et le noyau externe riche en fer et nickel.
La lithosphère et l’asthénosphère sont constituées de la même roche,
à quoi correspond alors la discontinuité située à 120 km de profondeur entre la lithosphère et l’asthénosphère ?
2) La limite lithosphère / asthénosphère.
- Le graphe issu des études expérimentales sur la péridotite montre l’évolution de la T° du manteau (géotherme) et la courbe de fusion (solidus) de la péridotite en fonction de la profondeur.
(Géotherme) : On constate que la T° de la péridotite augmente avec la profondeur : entre 150 et 300 km de profondeur, le géotherme est proche du solidus ; ce qui montre que la péridotite est presque fondue mais toujours solide. Dans cette zone, la péridotite est moins rigide que dans le reste du manteau, on parle de LVZ (Low velocity zone) : c’est l’asthénosphère.
Donc le manteau est constitué :
- de péridotites rigides parce que froides donc cassantes pour la lithosphère,
- de péridotites moins rigides parce que chaudes donc non cassables mais déformables pour l’asthénosphère .
3) La limite manteau supérieur / manteau inférieur.
On constate que :
- jusqu’à 400 km de profondeur, les ppx minéraux du manteau péridotitique sont l’olivine et lepyroxène,
- à partir de 400 km, ces minéraux changent de nature : on passe des spinelles et des grenats.
- vers 700 km de profondeur, une nouvelle structure, la pérovskite, remplace les structures précédentes et constitue tout le manteau inférieur.
La composition chimique de la péridotite ne change pas (ce sont toutes des péridotites), seule la densité varie, elle augmente avec la profondeur.
► Donc la différence entre la lithosphère et l’asthénosphère est due à la modification physique des minéraux de péridotite.
Les enveloppes sont séparées par des discontinuités physiques et / ou chimiques. La lithosphère se distingue de l’asthénosphère sous-jacente par un comportement rigide. La température, la pression et la masse volumique augmentent avec la profondeur.
Comment peut-on connaître la composition des roches en profondeur ? |
On sait grâce aux études sismiques que les enveloppes terrestres sont séparées par des discontinuités physiques et / ou chimiques. La lithosphère se distingue de l’asthénosphère sous-jacente par un comportement rigide. La température, la pression et la masse volumique augmentent avec la profondeur.
Comment peut-on connaître la composition des roches en profondeur grâce à l'étude des météorites ?
1) Formation des objets du système solaire.
La Terre , comme les autres planètes et autres objets du système solaire, s’est formée il y a 4,6 milliards d’années par accrétion, à partir d’une nébuleuse.
Accrétion : formation progressive d’un corps céleste par agglomération d’éléments (poussières, etc.) sous l’effet de la gravité. Les éléments formés sont de plus en plus gros et de plus en plus attractifs.
Toutefois, des objets rocheux ne se sont pas agglomérés : il s’agit des astéroïdes.
Astéroïdes = Objets célestes formés de roches, plus petits qu’une planète et de forme quelconque, gravitant normalement autour du soleil sur une orbite située entre celles de mars et de Jupiter. Ils quittent parfois leurs orbites.
Les astéroïdes qui tombent sur le sol d’une planète sont appelées météorites. Les météorites constituent donc des vestiges des roches constitutives de la Terre, accessibles à l’observation et à l’analyse chimique.
2) Les météorites et leur formation.
► Il existe différents types de météorites et leurs classifications sont diverses.
► Nous retiendrons que la plupart sont issues d’astéroïdes indifférenciés, il s’agit des chondrites alors que certaines représentent des fragments d’astéroïdes différenciés : les achondrites, sidérites et pallasites.
Quelles informations nous apportent ces météorites sur la Terre et sa formation ?
3) Apports des météorites sur la formation de la Terre.
a. Chondrites et composition globale de la Terre.
► Les météorites indifférenciées nous permettent de remonter les temps et de confirmer la première étape de la formation de la Terre : l’accrétion.
b. Météorites différenciées et formation de la Terre.
►Les pallasites sont particulièrement intéressantes car elles montrent une limite.
Par analogie avec les météorites différenciées, on admet aujourd’hui que la terre, de – 4,6 à – 4,4 Ma, s’est différenciée en plusieurs étapes :
- fusion d’un volume important de la Terre sous l’action de la chaleur interne et des impacts météoritiques très nombreux durant cette période ;
- le fer et le nickel, plus denses, migrent vers l’intérieur de la Terre (formation du noyau) ► sidérites ;
- les silicates fondus cristallisent et édifient le manteau et la croûte (^c en attestent les achondrites) ;
- une atmosphère se forme par dégazage.
Schéma de la «Terre solide différenciée », résultat d’une migration des éléments et d’une diminution de la T°
Conclusion :
Les matériaux du manteau profond et du noyau sont inaccessibles. On peut néanmoins, par des modèles et des raisonnements qui tiennent compte de la formation de la Terre à partir des chondrites, préciser leurs compositions.
La structure de la Terre, résulte d’une part de sa formation par accrétion de petits corps dont les météorites de type chondrite sont les vestiges, d’autre part de sa différenciation.
|
|
|
La lithosphère et la tectonique des plaques .
Corrigé |
La tectonique des plaques est une hypothèse, solidement étayée aujourd’hui, selon laquelle la partie superficielle de la Terre (lithosphère) est formée de plaques rigides d’une centaine de kilomètres d’épaisseur, en mouvements sur l’asthénosphère déformable. Ces plaques sont constituées d’une partie du manteau supérieur surmontée, suivant les cas, de croûte continentale ou océanique.
Comment les scientifiques ont-ils élaboré cette hypothèse ?
I. LES PREUVES DE LA TECTONIQUE DES PLAQUES.
A. Les fondements anciens de la tectonique des plaques.
On a longtemps pensé qu’un continent unique, la pangée, s’était segmenté en continents qui dérivaient à la surface du globe : c’est la théorie de la dérive des continents émise par Wegener en 1912.
Quelles ont été les preuves qui ont conforté cette idée ?
1) Des arguments géographiques.
En comparant les formes de certaines bordures de l’Amérique du sud et de l’Afrique, on s’aperçoit que si on les rapprochait, ces 2 continents s’emboîteraient comme les pièces d’un puzzle.
2) Des arguments géologiques.
L’âge des roches, l’orientation des chaînes de montagnes anciennes confortent l’hypothèse.
3) Des arguments paléoclimatiques.
On a retrouvé des traces d’évaporites, de charbons, témoins d’un climat chaud, datées d’il y a 200 MA sur divers continents éloignés actuellement. La distribution de ces roches s’explique bien si l’on admet que ces masses continentales étaient regroupées. Il en est de même pour d’autres roches sédimentaires.
4) Des arguments paléontologiques.
Des fossiles animaux et végétaux, de plus de 200 millions d’années ont été trouvés sur divers continents. S’il y avait eu présence d’un océan comme actuellement, il n’y aurait pas eu une telle dispersion d’espèces.
Tous ces arguments n’ont pas suffi à Wegener en 1912 pour imposer la théorie de la dérive des continents, par manque de preuve du moteur de cette dérive.
Il a fallu attendre les années 50 pour qu’elle soit reprise par d’autres chercheurs.
Problème : Comment peut-on prouver aujourd’hui la tectonique des plaques ?
B. Les preuves actuelles de la tectoniques des plaques.
1) Distribution géographique des volcans et des séismes.
La localisation des séismes est très inégale à la surface du globe. On observe des zones où pratiquement aucun séisme n'a lieu (zones asismiques): elles sont appelées plaques.
Elles sont délimitées entre elles par des zones où les séismes s'accumulent le long de bandes étroites :
- au « milieu » des océans : on parle de dorsales océaniques (Ex : Atlantique)
- à la limite océan-continent (pas toutes !) : ex Andes
- à l’intérieur des continents : au niveau des chaînes de montagnes (Ex : Himalaya)
2) Etude de la topographie des frontières de plaques.
La topographie (étymologiquement "écrire à propos d'un lieu") est la représentation graphique d'un lieu avec l'indication du relief.
a. Topographie d’une frontière océanique : la dorsale.
La dorsale est une zone de relief sous-marin, présentant une activité volcanique au niveau de la zone centrale appelée rift. Cette topographie suggère l’existence des mouvements de divergence à ce niveau.
b. Topographie d’une frontière océan / continent : une zone de subduction.
Certaines frontières océan-continent présentent une fosse profonde, des activités volcaniques et sismiques importantes qui suggèrent l’existence de mouvements de convergence.
c. Une frontière océanique plus discrète : la faille transformante.
Les dorsales océaniques sont recoupées par des failles transformantes qui les décalent ce qui suggère des mouvements de coulissement.
Les contours des bordures continentales (Wegener), la distribution géographique des volcans et des séismes et le relief de la Terre (topo), sont des signatures de la tectonique des plaques. La lithosphère est découpée en plaques d’épaisseur variable, peu déformables à l’exception de leurs limites.
Comment peut-on prouver et mesurer les mouvements des plaques ?
II. LES MOUVEMENTS DES PLAQUES.
A. Les données géologiques.
1) Apports de l’étude des points chauds.
Observations
En de nombreux endroits à la surface du globe on observe des alignements volcaniques intraplaques (qui ne sont pas situés au niveau d’une frontière entre 2 plaques).ex : les volcans de l’archipel de Hawaï sur la plaque Pacifique.
Ces alignements présentent les caractéristiques suivantes :
- un seul volcan est actif ; il est situé à l’extrémité de l’alignement
- plus on s’éloigne du volcan actif, plus l’âge des édifices volcaniques augmente.
Interprétation
Ces volcans proviennent de l’activité d’un point chaud, région fixe située dans le manteau (profond) qui envoie vers la surface « un panache » de magma perforant périodiquement la plaque sus-jacente. Sachant que les points chauds sont fixes et que leur activité est quasiment permanente, les alignements de volcans (ex Hawaï) traduisent le mouvement de la plaque sus-jacente au cours des temps géologiques.
La vitesse (absolue) du déplacement de la plaque est calculée à partir de l’âge du plus ancien volcan éteint et de sa distance par rapport au volcan actif actuel. |
 |
2) Apport de l’âge des fonds océaniques.
On mesure l’âge des sédiments déposés au dessus des basaltes. Ils présentent des âges symétriques par rapport à la dorsale n’excédant pas 180 Ma.
On en déduit que la fracturation de la Pangée a commencé il y a 180 Ma et depuis l ’expansion océanique a lieu de part et d’autre de la dorsale, de manière symétrique(en première approximation). La direction de l’expansion est donc perpendiculaire à la dorsale.
On peut mesurer la vitesse d'expansion de chaque côté. |
 |
3) Apports des anomalies magnétiques.
Origine des anomalies magnétiques :
Il existe un champ magnétique sur terre qui a son origine dans le noyau externe liquide. Il crée des lignes de champ orientées du pôle nord au pôle Sud. L’intensité du champ magnétique est variable selon l’endroit du globe où l’on se trouve, mais sa direction est fixe à un moment donné.
Le champ magnétique s’est inversé périodiquement au cours des temps géologiques.
Des roches volcaniques, comme le basalte, conservent les caractéristiques du champ magnétique terrestre qui régnait au moment de leur refroidissement. |
 |
| Observations et interprétation
:
Les anomalies magnétiques du plancher océanique basaltique :
- forment des bandes parallèles à l’axe de la dorsale
- présentent une alternance d’anomalies magnétiques positives et négatives (bandes de largeurs variables)
- sont disposées de manière symétrique par rapport à l’axe de la dorsale.
|
 |
Grâce aux anomalies magnétiques nous pouvons calculer la vitesse d’expansion océanique :
- océan Atlantique : 1 à 2 cm/an : (dorsale « lente »)
- océan Pacifique : 6 à 16 cm/an (dorsale « rapide »).
Différentes données géologiques (alignement des volcans de points chauds, âges des sédiments des fonds océaniques, anomalies magnétiques) permettent de reconstruire les directions et les vitesses des mouvements des plaques ainsi que leurs variations pour les 180 derniers millions d’années de l’histoire de la Terre .
Comment peut-on mesurer les vitesses aujourd’hui (en temps réel) ?
B. Mesure de la mobilité des plaques : utilisation du GPS.
Grâce aux techniques très modernes telles que les techniques de positionnement par satellites (GPS = Glogal Positioning Systeme) on peut mesurer la vitesse des déplacements des plaques et la distance parcourue avec une grande précision.
1) Principe de fonctionnement.
Les satellites possèdent des émetteurs qui envoient un signal ds les lg d’ondes qui traversent les nuages et qui est reçu par des balises de réception qui sont au sol.
Ces balises au sol reçoivent le signal d’au moins 4 satellites pour faire une mesure précise de position.
2) Tectonique des plaques et données GPS.
Les techniques de positionnement par satellites (GPS : Global Positioning System) mesurent les directions et vitesses des plaques sur des échelles de temps de quelques années.
Ces mesures pratiquement instantanées valident et affinent le modèle des déplacements globaux des plaques, fondé et construit sur des observations géologiques et géophysiques.
Conclusion :
Au cours du XX° siècle, la dérive des continents a laissé la place à la tectonique des plaques qui décrit et mesure les mouvements relatifs des plaques :
- des mouvements de divergence au niveau des dorsales océaniques où se forment les plaques ;
- des mouvements de convergence dans les zones de subduction et de collision où les plaques disparaissent ;
- des mouvements de coulissement le long des failles transformantes.
Reste-t-il des sceptiques ? |
Remarque : ressemble fort à une partie du sujet précédent !
On a longtemps pensé qu’un continent unique, la pangée, s’était segmenté en continents qui dérivaient à la surface du globe : c’est la théorie de la dérive des continents émise par Wegener en 1912. Cette théorie était basée sur des arguments géographiques, géologiques, paléoclimatiques et paléontologiques.
Toutefois, tous ces arguments n’ont pas suffi à Wegener en 1912 pour imposer la théorie de la dérive des continents, par manque de preuve du moteur de cette dérive.
Il a fallu attendre les années 50 pour qu’elle soit reprise par d’autres chercheurs. La tectonique des plaques est une hypothèse, solidement étayée aujourd’hui, selon laquelle la partie superficielle de la Terre (lithosphère) est formée de plaques rigides d’une centaine de kilomètres d’épaisseur, en mouvements sur l’asthénosphère déformable. Ces plaques sont constituées d’une partie du manteau supérieur surmontée, suivant les cas, de croûte continentale ou océanique.
Comment les scientifiques ont-ils étayé cette hypothèse ?
1) Des arguments géographiques.
Il s'agit d'un argument déjà apporté par Wegener. En comparant les formes de certaines bordures de l’Amérique du sud et de l’Afrique, on s’aperçoit que si on les rapprochait, ces 2 continents s’emboîteraient comme les pièces d’un puzzle.
2) Distribution géographique des volcans et des séismes.
La localisation des séismes est très inégale à la surface du globe. On observe des zones où pratiquement aucun séisme n'a lieu (zones asismiques): elles sont appelées plaques.
Elles sont délimitées entre elles par des zones où les séismes s'accumulent le long de bandes étroites :
- au « milieu » des océans : on parle de dorsales océaniques (Ex : Atlantique)
- à la limite océan-continent (pas toutes !) : ex Andes
- à l’intérieur des continents : au niveau des chaînes de montagnes (Ex : Himalaya)
3) Etude de la topographie des frontières de plaques. La topographie (étymologiquement "écrire à propos d'un lieu") est la représentation graphique d'un lieu avec l'indication du relief.
a. Topographie d’une frontière océanique : la dorsale.
La dorsale est une zone de relief sous-marin, présentant une activité volcanique au niveau de la zone centrale appelée rift. Cette topographie suggère l’existence des mouvements de divergence à ce niveau.
b. Topographie d’une frontière océan / continent : une zone de subduction.
Certaines frontières océan-continent présentent une fosse profonde, des activités volcaniques et sismiques importantes qui suggèrent l’existence de mouvements de convergence.
c. Une frontière océanique plus discrète : la faille transformante.
Les dorsales océaniques sont recoupées par des failles transformantes qui les décalent ce qui suggère des mouvements de coulissement.
Les contours des bordures continentales (Wegener), la distribution géographique des volcans et des séismes et le relief de la Terre (topo), sont des signatures de la tectonique des plaques. La lithosphère est découpée en plaques d’épaisseur variable, peu déformables à l’exception de leurs limites.
Comment peut-on prouver et mesurer les mouvements des plaques ?
|
I. Les dorsales océaniques.
A. Mouvements de divergence.
1) L'accrétion océanique.
2) Preuves de l'accrétion : paléomagnétisme, volcanisme de point chaud, âges des sédiments au contact des basaltes.
B. Formation des plaques.
II. Les marges actives.
A. Mouvements de convergence :
1) Subduction : volcanisme et séismes (profondeur des foyers).
2) Collision : formation d'une chaîne de montagnes.
B. Disparition de plaques.
III. Les failles transformantes.
A. Mouvements de coulissement.
B. Ni gain ni perte.
|
Remarque : ressemble fort à une partie du sujet n°1 !
Les contours des bordures continentales (Wegener), la distribution géographique des volcans et des séismes et le relief de la Terre (topo), sont des signatures de la tectonique des plaques. La lithosphère est découpée en plaques d’épaisseur variable, peu déformables à l’exception de leurs limites.
Comment peut-on reconstruire les directions et les vitesses des mouvements des plaques ?
I. Les données géologiques.
1) Apports de l’étude des points chauds.
Observations
En de nombreux endroits à la surface du globe on observe des alignements volcaniques intraplaques (qui ne sont pas situés au niveau d’une frontière entre 2 plaques).ex : les volcans de l’archipel de Hawaï sur la plaque Pacifique.
Ces alignements présentent les caractéristiques suivantes :
- un seul volcan est actif ; il est situé à l’extrémité de l’alignement
- plus on s’éloigne du volcan actif, plus l’âge des édifices volcaniques augmente.
Interprétation
Ces volcans proviennent de l’activité d’un point chaud, région fixe située dans le manteau (profond) qui envoie vers la surface « un panache » de magma perforant périodiquement la plaque sus-jacente. Sachant que les points chauds sont fixes et que leur activité est quasiment permanente, les alignements de volcans (ex Hawaï) traduisent le mouvement de la plaque sus-jacente au cours des temps géologiques.
La vitesse (absolue) du déplacement de la plaque est calculée à partir de l’âge du plus ancien volcan éteint et de sa distance par rapport au volcan actif actuel. |
|
2) Apport de l’âge des fonds océaniques.
On mesure l’âge des sédiments déposés au dessus des basaltes. Ils présentent des âges symétriques par rapport à la dorsale n’excédant pas 180 Ma.
On en déduit que la fracturation de la Pangée a commencé il y a 180 Ma et depuis l ’expansion océanique a lieu de part et d’autre de la dorsale, de manière symétrique(en première approximation). La direction de l’expansion est donc perpendiculaire à la dorsale.
On peut mesurer la vitesse d'expansion de chaque côté. |
|
3) Apports des anomalies magnétiques.
Origine des anomalies magnétiques :
Il existe un champ magnétique sur terre qui a son origine dans le noyau externe liquide. Il crée des lignes de champ orientées du pôle nord au pôle Sud. L’intensité du champ magnétique est variable selon l’endroit du globe où l’on se trouve, mais sa direction est fixe à un moment donné.
Le champ magnétique s’est inversé périodiquement au cours des temps géologiques.
Des roches volcaniques, comme le basalte, conservent les caractéristiques du champ magnétique terrestre qui régnait au moment de leur refroidissement. |
|
| Observations et interprétation :
Les anomalies magnétiques du plancher océanique basaltique :
- forment des bandes parallèles à l’axe de la dorsale
- présentent une alternance d’anomalies magnétiques positives et négatives (bandes de largeurs variables)
- sont disposées de manière symétrique par rapport à l’axe de la dorsale.
|
|
Grâce aux anomalies magnétiques nous pouvons calculer la vitesse d’expansion océanique :
- océan Atlantique : 1 à 2 cm/an : (dorsale « lente »)
- océan Pacifique : 6 à 16 cm/an (dorsale « rapide »).
Différentes données géologiques (alignement des volcans de points chauds, âges des sédiments des fonds océaniques, anomalies magnétiques) permettent de reconstruire les directions et les vitesses des mouvements des plaques ainsi que leurs variations pour les 180 derniers millions d’années de l’histoire de la Terre .
Comment peut-on mesurer les vitesses aujourd’hui (en temps réel) ?
II. Mesure de la mobilité des plaques : utilisation du GPS.
Grâce aux techniques très modernes telles que les techniques de positionnement par satellites (GPS = Glogal Positioning Systeme) on peut mesurer la vitesse des déplacements des plaques et la distance parcourue avec une grande précision.
1) Principe de fonctionnement.
Les satellites possèdent des émetteurs qui envoient un signal ds les lg d’ondes qui traversent les nuages et qui est reçu par des balises de réception qui sont au sol.
Ces balises au sol reçoivent le signal d’au moins 4 satellites pour faire une mesure précise de position.
2) Tectonique des plaques et données GPS.
Les techniques de positionnement par satellites (GPS : Global Positioning System) mesurent les directions et vitesses des plaques sur des échelles de temps de quelques années.
Ces mesures pratiquement instantanées valident et affinent le modèle des déplacements globaux des plaques, fondé et construit sur des observations géologiques et géophysiques.
Conclusion :
Au cours du XX° siècle, la dérive des continents a laissé la place à la tectonique des plaques qui décrit et mesure les mouvements relatifs des plaques :
- des mouvements de divergence au niveau des dorsales océaniques où se forment les plaques ;
- des mouvements de convergence dans les zones de subduction et de collision où les plaques disparaissent ;
- des mouvements de coulissement le long des failles transformantes.
Grâce aux données géologiques et au GPS, on peut aujourd'hui mesurer les mouvements des plaques.
Reste-t-il des sceptiques ? |
|
| |
| Divergence et phénomènes liés.
Corrigés |
Sujet 1 :
La tectonique des plaques est aujourd'hui largement prouvée, notamment grâce aux techniques de positionnement par satellites (GPS : Global Positioning System) qui mesurent les directions et vitesses des plaques sur des échelles de temps de quelques années.
Comment la Pangée s’est-elle fracturée et a-t-elle abouti à la distribution actuelle des plaques ?
En étudiant tout d'abord une modélisation en plâtre, nous montrerons que l'on peut mettre en évidence une partie des indices géologiques prouvant l'expansion. Nous dresserons enfin un bilan des étapes "du continent à l'océan".
I. Modélisation de l'expansion.
A. Réalisation du modèle.
 |
Nous créons un modèle dans un aquarium (1) percé de 2 trous latéraux par lesquels passent des planches (2) qui maintiennent le plâtre et que l'on peut tirer grâce aux "manettes" (3).
Cinq couches de plâtre (4) colorées à la craie écrasée ont été déposées puis compactées. |
On tire alors sur les manette pour modéliser l'expansion.
B. Résultats obtenus.
Les couches se "fissurent" : des failles se forment. Au centre on observe un effondrement. Globalement les terrains ont "gagné" en longueur mais se sont amincis.
Ces résultats correspondent-ils aux preuves géologiques de l'expansion ?
II. Observations géologiques.
A. En concordance avec le modèle.
La déchirure continentale se caractérise par un amincissement crustal, la formation de failles normales et donc de blocs basculés : on parle de rifting continental,
Les observations géologiques confirment les données du modèle et les complètent. En effet, un rift continental est constitué de bassins sédimentaires comblés par des sédiments fluviatiles, lacustres ou détritiques. Des roches anciennes, sédimentaires, en témoignent.
Une mer peu profonde s’installe alors dans le fossé ainsi formé, des sédiments peuvent alors se déposer sur les « marches » constituées par les blocs basculés. Les roches sédimentaires qui en résultent sont principalement des évaporites, liées à une intense évaporation, comme le gypse (forment notamment les roses des sables).
Lorsqu’une mer plus profonde s’installe, des roches comme le calcaire, les marnes se forment.
B. En complément du modèle.
Le modèle en sable permet de visualiser, de comprendre un phénomène long mais il est incomplet (pas de volcanisme…) et ne reproduit pas exactement la réalité : les roches ne sont pas du sable !
Les rifts continentaux sont également le siège d’une activité volcanique importante et d’une activité sismique.
D'après ces informations, quelles sont les différentes étapes de la formation d'un océan ?
III. De la Pangée à la distribution actuelle des plaques.
A. Stade de distension ou rifting continental.
observable en Alsace, en Limagne et dans la région des grands lacs africains. Il y a création de fossés, au centre de vastes bombements topographiques présentant un flux géothermique supérieur à la moyenne, un volcanisme basaltique, une sismicité élevée, une sédimentation épaisse et souvent évaporitique.
B. Stade océan étroit.
Il y a eu création d'un fond océanique, mais la faible largeur de l'océan interdit le renouvellement de l'eau profonde par les courants, ce qui entraîne leur stagnation, empêche leur oxygénation et engendre un milieu où la matière organique se conserve (boues noires). Ce stade est aujourd'hui en partie illustré par la mer Rouge.
C. Stade océan large.
Comme l'actuel Atlantique, la circulation des eaux entraînées par les courants y est aisée et, sur les fonds, l'oxygène se renouvelle : la matière organique s'y oxyde et disparaît. La répartition de la sédimentation est alors essentiellement gouvernée par 3 paramètres : la quantité des apports détritiques, la productivité biologique du milieu océanique, l'éloignement de la dorsale, qui conditionne l'âge du fond océanique et donc sa profondeur.
Conclusion : Failles normales, sédimentation continentale et volcanisme sont trois marqueurs géologiques de la rupture de la lithosphère continentale. Le rift est ensuite envahi par la mer puis un océan se crée quand le magma forme le plancher océanique.
Les observations géologiques et modèles construits à partir de ces dernières ont permis de comprendre des phénomènes que l'on ne peut appréhender à l'échelle de la vie humaine.
Que deviendra ensuite cet océan ?
|
Sujet 2 :
Une très longue chaîne de montagnes parcourt le globe, discrètement puisque sous les océans. A son niveau, des basaltes en coussins se mettent en place en surface mais aussi de gabbros en profondeur.
Comment se forme un magma basaltique et comment ce dernier peut-il donner naissance à des basaltes mais aussi des gabbros ?
Nous répondrons à cette problématique en présentant la fusion partielle des péridotites du manteau puis les causes et conséquences de cette fusion partielle. Enfin nous verrons comment le refroidissement conduit à des roches de textures différentes (basaltes/gabbros).
I. Fusion partielle des péridotites et formation du magma.
A. Informations apportées par la sismique réflexion.
Sous la dorsale, on remarque :
- un bombement du Moho matérialisant une remontée du manteau et donc un amincissement crustal ;
- une interruption du Moho sous l’axe de la dorsale montrant le lieu de remontée des liquides issus de la fusion de la péridotite du manteau sous jacent ;
- un espace retardant les ondes sismiques : la chambre magmatique, lieu contenant (de collecte) du magma.
B. Quantité de magma formée.
En ce qui concerne la production de magma mondiale, elle peut être estimée de la manière suivante :
On considère en moyenne une expansion océanique de 5cm/an (=5. 10 -5 km/an), avec une production de croûte océanique de 7 km d'épaisseur sur les 60 000 km de dorsale, ce qui donne un volume de roches magmatiques (1/4 de basalte, 3/4 de gabbro) produit de 7 x 60 000 x 5 x 10 -5 soit 21 km 3.
Ceci nous démontre qu’une très petite partie du globe fond.
Les dorsales sont le siège d’une production importante de magma : de l’ordre de 20 km 3 par an.
Comment expliquer cette fusion ?
II. Causes et conséquences de la fusion partielle.
A. Conditions de fusion de la péridotite.
A une profondeur de 100 km à la verticale d’une dorsale, la température de la péridotite asthénosphérique est de 1300°C. Pour expliquer la fusion de cette péridotite, il faut envisager une remontée sans perte de chaleur mais avec diminution de pression : on parle de décompression adiabatique.
La péridotite ne fond pas totalement (liquidus non franchi) mais en partie : on parle de fusion partielle.
Les magmas sont issus de la fusion partielle des péridotites du manteau induite par décompression. Ils sont de nature basaltique.
Quelle proportion de la péridotite fond ? Quelles en sont les conséquences ?
B. Fusion partielle de la péridotite.
Seulement 5 à 15 % de la péridotite fond.

La péridotite a une composition chimique différente de celle du magma basaltique parce que la fusion de la péridotite est partielle. En effet, lors de la fusion partielle, une différenciation chimique se produit.
La péridotite est composée d’olivine, de pyroxène (minéral ferro-magnésien) et de plagioclase (minéral alumino-calcique).
Le plagioclase fond en premier. Le liquide magmatique aura donc une composition chimique plus riche en calcium et aluminium que la péridotite initiale. La péridotite résiduelle sera enrichie en olivine et pyroxène.
En définitive, la composition chimique du magma basaltique dépend en premier du taux de fusion partielle des péridotites.
La fusion partielle donne au magma basaltique une composition chimique différente de celle de la roche source.
III. Refroidissement du magma et formation des roches de la croûte océanique.
A. Rappel : comparaison gabbro – basalte. Le gabbro et le basalte présentent la même composition minéralogique mais une structure différente :
- le gabbro est totalement cristallisé : il a une structure grenue ;
- le basalte présente quelques cristaux observables à l’œil nu (phénocristaux), de nombreux microlites et une matrice non cristallisée (du verre) : il a une structure microlitique.
Comment peut-on expliquer cette différence de structure ?
B. L’origine de la différence de structure entre basalte et gabbro. Hypothèse : la vitesse de cristallisation conditionne la structure de la roche.
La cristallisation expérimentale.
- La cristallisation lente de la vanilline provoque la formation de gros cristaux jointifs : la structure est grenue.
- La cristallisation rapide (sur un bloc de glace) de la vanilline provoque la formation de petits cristaux non jointifs, une partie de la lame n’est pas cristallisée : la structure est microlitique.
L’hypothèse est validée : la structure d’une roche dépend de sa vitesse de cristallisation.
De plus on remarque que les minéraux ne se forment pas en même temps : on dit qu’il y a cristallisation fractionnée.
Def : formation de minéraux différents à des moments successifs dans un magma qui se refroidit. Lorsque la température du magma décroît, les minéraux qui cristallisent à haute température se forment les 1° (ici olivine et pyroxène). Ces minéraux étant riches en Fe, Mg et Ca, le magma résiduel est appauvri en ces éléments et corrélativement enrichi en Si, Al, K, Na.
La cristallisation fractionnée explique en partie la composition chimique différente entre les roches magmatiques.
Le refroidissement plus ou moins rapide des magmas conduit à des roches de textures différentes (basaltes / gabbros).
Conclusion : fusion partielle puis cristallisation fractionnée permettent de former les magmas puis les roches de la lithosphère océanique et donc contribuent à l’expansion océanique.
Comment l’océan va-t-il ensuite évoluer ? |
Sujet 3 :
Une marge passive est la zone de transition entre croûte continentale et croûte océanique fo rmant la bordure stable d’un continent, sans manifestation sismique ni volcanique.
Comment l’étude géologique des marges passives permet de déduire qu'elles ont enregistré l'histoire précoce de la rupture continentale et de l'océanisation ?
Nous étudierons successivement les caractéristiques morphologiques, tectoniques puis sédimentaires des marges passives pour résoudre la problématique.
I. Les caractéristiques morphologiques des marges passives.
En partant du continent vers l’océan, la topographie (étude des reliefs) d’une marge passive montre :
- le plateau continental de pente très douce ( 200 m de profondeur) et large de70 à 80 km en moyenne ;
- le talus va de 200m à 3000m de profondeur avec une pente très prononcée ;
- le glacis entre 3000 et 5000m qui se raccorde à la plaine abyssale pratiquement horizontale
II. Les caractéristiques tectoniques des marges passives. Le profil sismique montre une croûte continentale amincie depuis la ligne de côte ( 30 km) jusqu’au glacis où elle se raccorde à la croûte océanique (7 à 10 km).Cette croûte est fracturée par des failles normales inactives actuellement délimitant des blocs basculés vers le continent (affaissement et glissement vers l’océan, basculement vers cont). Ces déplacements caractérisent un étirement de la zone qui s’est allongée de 30 à 40 %.
La croûte fracturée est recouverte de sédiments.
Quelles informations nous apporte l’étude des roches sédimentaires des marges passives ?
III. Les caractéristiques sédimentaires des marges passives.
La lithosphère rigide est en équilibre (hydrostatique) sur l’asthénosphère moins rigide : c’est le principe de l’isostasie. Un alourdissement est responsable de l’enfoncement : on parle de subsidence c'est-à-dire un affaissement progressif de la région. La subsidence assure une sédimentation importante.
On trouve au niveau d’une marge passive 3 types de formations sédimentaires :
- Les formations sédimentaires pré-rift, déposées avant la fracturation du continent, présentent une disposition en strates parallèles à la surface des blocs basculés, elles sont recoupées et décalées par les failles ce qui prouve leur antériorité.
- Les formations sédimentaires de type syn-rift sont affectés par les failles, donc elles sont contemporaines de la formation des failles. Elles témoignent du stade de distension ou rifting continental (aujourd’hui observable en Alsace, en Limagne et dans la région des grands lacs africains) . Il s’agit de formations évaporitiques.
- Les formations post-rift recouvrent les failles, les strates ne sont pas affectées car elles se sont mises en place après la phase d’ouverture, au stade océan étroit ou postérieurement. Les boues noires, riches en hydrocarbures témoignent du stade océan étroit. Les calcaires, marnes qui se situent au dessus témoignent du stade océan large.
Les formations syn-rift datent du Crétacé < et les formations post-rift du crétacé >. Les sédiments syn-rift sont contemporains des failles, la marge a donc pris naissance au crétacé <. L’océanisation révélée par la présence de croûte océanique est datée de 100 Ma.
Conclusion (et notion) : Les marges passives des continents sont structurées par des failles normales et sont le siège d’une sédimentation importante. L’activité des failles normales, héritage de rifts continentaux, témoigne de l’amincissement de la lithosphère et de sa subsidence. Les marges passives ont enregistré l’histoire précoce de la rupture continentale et de l’océanisation.
|
|
|
La machinerie thermique de la Terre.
|











 créés par
créés par